Nature Cell Biology丨一項重要的細胞重編程研究成果發(fā)表
2018年3月12日,國際著名期刊《Nature Cell Biology》(影響因子20.06)刊登了中國科學院廣州生物醫(yī)藥與健康研究院Miguel A. Esteban教授團隊題為NCoR/SMRT co-repressors cooperate with c-MYC to create an epigenetic barrier to somatic cell reprogramming的重磅成果,研究發(fā)現(xiàn)NCoR/SMRT輔阻遏物可結合多能性位點,與c-MYC作用最終導致重編程中的表觀遺傳障礙,該研究對揭示這一類輔阻遏物在重編程中關鍵角色有重要意義。
OSKM與轉錄共同調節(jié)因子之間互作誘導機制的難解之謎?
眾所周知,體細胞可以通過外源因子OSKM (OCT4、SOX2、KLF4和c-MYC) 來重編程為誘導多能干細胞(iPSCs)。重編程開始時,外源性OSKM與整個基因組的DNA結合并誘導連續(xù)的染色質重組,從而激活整個多能性基因網(wǎng)絡。然而,OSKM不能單獨運作,需要共同調節(jié)因子來修飾局部表觀遺傳環(huán)境。但目前尚不清楚OSKM和不同轉錄共同調節(jié)因子(共激活因子和輔阻遏物)之間是如何相互作用或相互拮抗以誘導多能性狀態(tài)。
Miguel A. Esteban教授團隊探索了兩個已知的輔阻遏物核受體輔助抑制因子(NCoR)和視黃酸及甲狀腺激素受體沉默中介蛋白(SMRT)的功能,證明了NCoR/SMRT輔阻遏物可與多能性位點結合,從而利用4個Yamanaka因子(OCT4、SOX2、KLF4和c-MYC)創(chuàng)建一個重編程障礙,發(fā)現(xiàn)抑制NCoR/SMRT就可顯著提高重新編程效率和動力學。NCoR/SMRT復合物的核心表觀遺傳亞基-組蛋白去乙酰化酶3(HDAC3)可通過誘導多能性位點上的組蛋白去乙酰化而有助于NCoR/SMRT發(fā)揮作用。在Yamanaka因子中,主要由c-MYC促進NCoR/SMRT-HDAC3在基因組位點的募集。
最后,研究人員描述了c-MYC是如何對重編程的早期階段有益,而對后期階段有害。總體而言,本研究發(fā)現(xiàn)了NCoR/SMRT輔阻遏物在重編程中的作用,并在提出了c-MYC的雙重功能機制。
心臟發(fā)育是一個多步驟的過程,取決于對網(wǎng)絡的精確控制,包括多種心臟特異性轉錄因子、染色質修飾因子和信號通路。T-box轉錄因子Eomes缺失顯著損害中內胚層分化,包括心臟中胚層特化。然而,Eomes如何參與特定的心肌分化尚不清楚。先前的研究表明Eomes通過直接結合Mesp1基因的啟動子來誘導Mesp1表達。但是Eomes在心臟中胚層特化期間誘導Mesp1表達的精確調控機制仍然不清楚。
論文中的6項重要研究發(fā)現(xiàn)
1) NCoR/SMRT輔阻遏物可導致OSKM重編程障礙
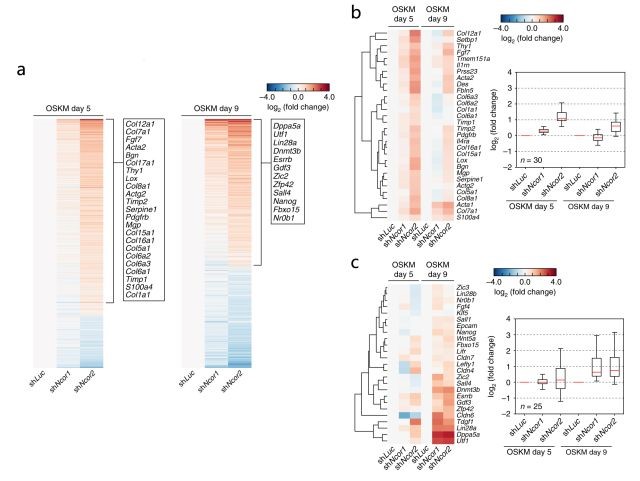
圖1
通過RT–qPCR、shRNA敲降實驗及熒光素酶報告實驗等證明抑制Ncor1(編碼NCoR)和Ncor2(編碼SMRT),或者抑制NCoR/SMRT可增強OSKM重編程。
那么,NCoR和SMRT是如何阻礙重編程的呢?通過對OSKM重編程第5天和第9天的Ncor1/2缺失細胞進行RNA-seq(由銳博生物提供),發(fā)現(xiàn)Ncor1/2的敲降使得許多MEF富集的體細胞基因在第5天時被上調,在第9天時變得不明顯;而多能性相關基因在第9天時被上調(圖1)。
因此,OSKM重編程中,抑制Ncor1/2首先會導致體細胞基因抑制中的瞬時缺陷,然后導致多能性基因的加速及更有效的激活。
2) HDAC3對于NCoR/SMRT輔阻遏物損害的重編程至關重要
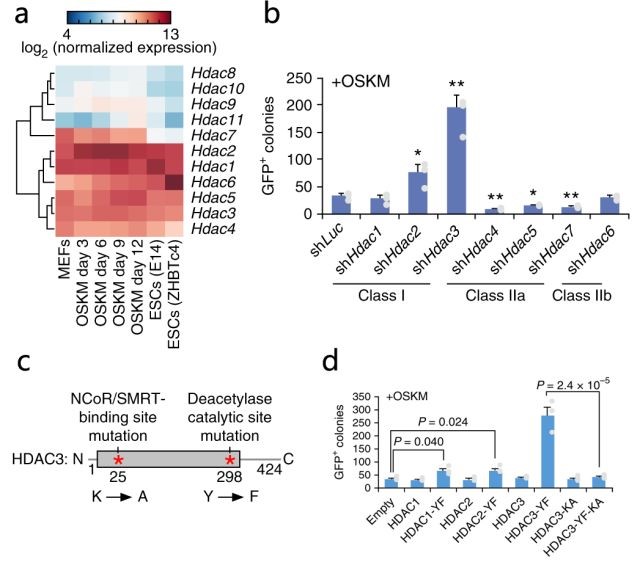
圖2
HDAC3作為NCoR/SMRT復合物的核心表觀遺傳亞基,負責復合物的去乙酰化酶活性。研究人員通過RNA-seq分析(由銳博生物提供)及敲降實驗發(fā)現(xiàn)Hdac3在OSKM介導的重編程中表達,且敲降Hdac3顯著增加重編程(圖2ab)。
隨后構建了HDAC3去乙酰化酶無效突變體(HDAC3-YF)、HDAC3雙突變體(HDAC3-YF-KA,與NCoR/SMRT相互作用的能力降低),發(fā)現(xiàn)HDAC3-YF能有效增強重編程,但這種增強作用在HDAC3-YF-KA突變體中消失,表明HDAC3-YF需要與NCoR/SMRT相互作用才能達到增強重編程的作用(圖2cd)。因此推斷HDAC3可介導NCoR/SMRT損害OSKM重編程,并且該功能需要去乙酰化酶活性。
3) HDAC3在多能性基因組位點誘導組蛋白去乙酰化

圖3
HDAC3是如何損害OSKM重編程呢?通過免疫印跡分析發(fā)現(xiàn)OSKM重編程過程中組蛋白乙酰化水平增加,而NCoR/SMRT缺失或HDAC3-YF過表達導致其水平不再增加。然后在OSKM重編程的第5/9/13天針對H3K27ac進行ChIP–seq以驗證其對組蛋白乙酰化的影響(由銳博生物提供),發(fā)現(xiàn)大多數(shù)H3K27乙酰化(H3K27ac)位于轉錄起始位點(TSS)附近,有4個多能性位點(Sall4、Utf1、Nanog和Zic2))在第9/13天時顯示H3K27ac的增加,此外,OSKM和HDAC3-YF重編程細胞的體細胞基因座中H3K27ac的水平略高,但只在少數(shù)情況下且僅在第5天(圖3)。表明,HDAC3作為NCoR/SMRT復合物的一部分,通過限制性位點(包括多能性位點)誘導組蛋白去乙酰化而損害OSKM重編程。
4) NCoR/SMRT在重編程中可被募集到多能性位點
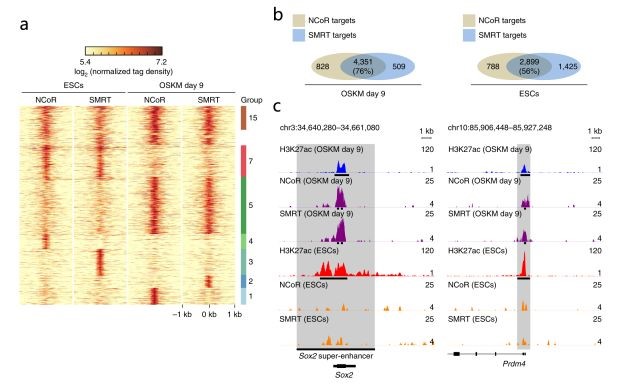
圖4
NCoR/SMRT是否會被募集到多能性位點以通過HDAC3誘導組蛋白去乙酰化?研究人員對OSKM重編程第9天的serum+Vc樣本進行ChIP–seq(由銳博生物提供),發(fā)現(xiàn)很大一部分NCoR/SMRT結合峰是重編程(76%)和ESCs(56%)共有的,主要位于TSS附近(圖4ab)。進一步分析發(fā)現(xiàn)NCoR/SMRT僅在重編程過程中被募集到多能性位點Sox2和Prdm4,且這些位點具有更低水平的H3K27ac(圖4c),與NCoR/SMRT通過HDAC3損害重編程結果一致。
5) NCoR/SMRT–HDAC3通過c-MYC被募集到多能性位點

圖5
通過免疫共沉淀發(fā)現(xiàn)c-MYC與NCoR/SMRT-HDAC3復合體之間相互作用。ChIP-seq數(shù)據(jù)分析發(fā)現(xiàn)pre-iPSCs中NCoR/SMRT與c-MYC有最強的重疊(72%的NCoR/SMRT峰),其次是KLF4(44%)和OCT4/SOX2(~5%),但大多數(shù)NCoR/SMRT與KLF4的重疊位點被c-MYC共同結合(圖5a-d)。OSKM重編程中NCoR/SMRT與OCT4、SOX2及c-MYC同樣有很強的重疊,且大部分位點都是共有的(圖5e-g)。因此,c-MYC主要幫助將NCoR/SMRT募集到基因組位點(包括多能性位點)以誘導重編程的負效應。
6) 外源性c-MYC在重編程中的雙重作用

圖6
為了進一步評估c-MYC和NCoR/SMRT在抑制多能性基因中的關系,構建了DsRed慢病毒報告基因載體(可通過多能性轉錄因子的結合直接激活),發(fā)現(xiàn)單獨的OCT4、SOX2和KLF4過表達可適當激活報告基因,它們結合后(OSK)的激活效應更強;而單獨的c-MYC沒有激活效應,結合OSK會損害報告基因活性的增加。Ncor1/2敲降部分緩解了c-MYC對報告基因的負效應(fig8bc)。研究表明通過c-MYC將NCoR/SMRT輔阻遏物募集到多能性位點是重編程的負效應力,但將c-MYC添加到OSK中又可增強重編程效率。后續(xù)實驗證明了外源性c-MYC抑制多能性位點對于通過募集NCoR/SMRT-HDAC3進行重編程的后期階段是不利的。


