銳博生物RICK技術最新成果在《Nature Methods》發(fā)表!
全面揭示新生RNA相互作用的神秘面紗!
2018年2月12日由中國科學院廣州生物醫(yī)藥與健康研究院Miguel A. Esteban(米格爾·埃斯特班)、張必良教授和鮑習琛副研究員領銜的科研團隊在《Nature Methods》雜志在線發(fā)表了題為“Capturing the interactome of newly transcribed RNA”的研究論文,利用新設計的分離RNA結合蛋白的RICK技術(Newly Transcribed RNA interactomeusing click chemistry)研究人員系統(tǒng)地分離了包括非polyA尾RNA和新生RNA在內(nèi)的一系列RNA分子及其結合蛋白。進一步分析發(fā)現(xiàn),細胞分裂調(diào)控因子作為新的RNA結合蛋白,存在結合非polyA尾RNA的潛能。研究人員縮短RNA標記的時間,成功地在細胞中分離了新生RNA結合蛋白。這些蛋白與新生RNA的轉(zhuǎn)錄及后續(xù)的加工、剪切等調(diào)控密切相關。由于RICK很容易修改,例如標記時間或轉(zhuǎn)錄干擾的變化,所以它適用于不同的細胞類型和條件將有助于實現(xiàn)RNA結合蛋白質(zhì)組的真實維度。這一技術的應用將有助于深入地剖析RNA結合蛋白的作用機制,也是細胞命運轉(zhuǎn)變過程中分析RNA蛋白相互作用的重要工具。
摘? ?要?
將EU(5-ethynyluridine)標記的新轉(zhuǎn)錄RNAs與結合蛋白的表征相結合的方法稱為RICK(利用點擊化學捕獲新轉(zhuǎn)錄的RNA相互作用組),該方法可系統(tǒng)地捕獲與許多各種不同RNAs(包括新生RNAs和傳統(tǒng)上被忽略的非polyA RNAs)結合的蛋白。RICK已經(jīng)確定了對非polyA RNAs具有優(yōu)先親和力的其他新型RNA結合蛋白中的有絲分裂調(diào)節(jié)因子,揭示了代謝酶/因子與新生RNAs之間的聯(lián)系,并擴展了小鼠胚胎干細胞(mESCs)的已知RNA結合蛋白質(zhì)組。RICK將促進對不同細胞和系統(tǒng)中總RNA結合蛋白質(zhì)組的深入研究。
研究背景
目前,系統(tǒng)描述RNA結合蛋白質(zhì)組(或RNA相互作用組)的方法主要是基于用oligo(dT)包被的磁珠捕獲聚腺苷酸化(polyA)RNA,主要是mRNA。然而,polyA尾僅在其加工過程中被添加到新轉(zhuǎn)錄的RNAs中成為成熟的形式,而一些成熟的mRNAs是非polyA或bimorphic(二態(tài))。而且,成熟的非polyA RNA種類包含全部轉(zhuǎn)錄序列相當大的一部分。考慮可以捕獲與所有類型RNAs相互作用的RNA結合蛋白(RBPs)的方法學不僅能夠擴展對RNA相互作用組的現(xiàn)有觀點,而且還有助于理解非polyA RNAs在生理學和疾病中的功能。研究者基于使用點擊反應將EU標記的RNAs和生物素進行鏈接,開發(fā)了一種通用方法來捕獲新轉(zhuǎn)錄的RNA相互作用組。
研究路線??
1、設計RICK技術,用于捕獲新轉(zhuǎn)錄的RNA相互作用組:EU、凝膠電泳、銀染、pull-down、western blotting等
2、鑒定RICK捕獲的RNA種類:RNA-seq、RT-qPCR等
3、RICK分離的蛋白質(zhì)的特征及功能分析:LC-MS/MS、western blotting、GO及KEGG通路分析等
4、RICK鑒定優(yōu)先結合非polyA RNAs的蛋白質(zhì):RT-PCR、LC-MS/MS、western blotting等
5、與METTL1和CDK1相互作用的RNAs的表征:PAR-CLIP測序、RIP-qPCR等
6、RICK捕獲新生RNA相互作用組:EU、GO及KEGG通路分析等
7、RICK捕獲mESCs的總RNA相互作用組:EU、LC-MS/MS等
研究結果??
1)??使用RICK捕獲新轉(zhuǎn)錄的RNA相互作用組
研究者首先用EU標記HeLa細胞中的RNA(16h),然后使用254nm UV將RNA與蛋白進行交聯(lián),并使用點擊反應對EU進行生物素化。接下來,通過鏈霉親和素偶聯(lián)的磁珠提取EU標記的RNA-蛋白質(zhì)復合物(圖1)。凝膠電泳和銀染確認RICK成功分離出與EU標記的RNA直接相互作用的蛋白質(zhì)。當樣品未交聯(lián)或與RNase共處理時,蛋白質(zhì)pull-down信號丟失,證實pull-down的特異性。western blotting驗證了RICK對已知RBPs的捕獲。因此,使用新建立的RICK可特異性分離與新轉(zhuǎn)錄RNA相互作用的蛋白質(zhì)。

圖? 1
2)??測定RICK捕獲的RNA種類
對RICK pull-down樣本進行RNA-seq,發(fā)現(xiàn)與oligo(dT)捕獲數(shù)據(jù)存在明顯差異(圖2a)。接著,研究了RICK樣本中3種相關的非polyA RNA種類:環(huán)狀RNAs(circRNAs)、近端啟動子RNAs(ppRNAs)和增強子RNAs(eRNAs)。與oligo(dT)捕獲數(shù)據(jù)相比,在RICK樣本中觀察到6199個反向剪接事件(暗示為circRNAs),其中828個與circBase數(shù)據(jù)集重疊。另外,轉(zhuǎn)錄暫停基因(RNA Pol II Traveling Ratio/TR>4)在TSS周圍顯示出更高的RNA-seq信號積累,表明僅通過RICK可有效分離ppRNAs,而對于非暫停基因(TR<4)則差異很小(圖2bc)。此外,通過RICK樣品中假定的增強子區(qū)域還觀察到強烈富集的RNA-seq信號(提示為eRNAs),而不是oligo(dT)捕獲(圖2d)。定量RT-PCR驗證RICK提取的非polyA RNAs,發(fā)現(xiàn)oligo(dT)捕獲樣品中相同的RNA較少富集或不存在(圖2e)。研究顯示,RICK成功地分離了不限于polyA RNAs的各種各樣RNA種類,表明它也可以富集oligo(dT)捕獲方法不能分離的RBPs。
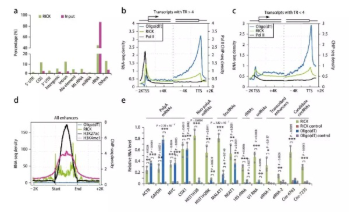
圖? 2
3)???表征RICK分離的蛋白質(zhì)
對RICK分離的蛋白進行LC-MS/MS分析,發(fā)現(xiàn)1353種蛋白質(zhì),其中720種高置信度蛋白,633種低置信度蛋白。與oligo(dT)捕獲數(shù)據(jù)相比,RICK樣本中存在344種RICK獨有的RBPs。
然后,在EU存在的情況下進行oligo(dT)捕獲,以評估其摻入是否影響RNA-蛋白質(zhì)相互作用。聚類分析發(fā)現(xiàn)EU+和EU- oligo(dT)捕獲樣品彼此之間高度相關,但與RICK樣品相關性不大(圖3a)。EU+ oligo(dT)捕獲樣品有469種高置信度蛋白,僅有2種在344種RICK獨有的RBPs之中。
將344種RICK獨有的RBPs與3個報道的人oligo(dT)捕獲研究進行比較,發(fā)現(xiàn)大多數(shù)這些蛋白質(zhì)(295,85.8%)對于RICK來說確實是獨特的(圖3b)。Western blotting證實了幾個RICK獨有的RBPs(如有絲分裂調(diào)節(jié)因子CDK1和m7G甲基轉(zhuǎn)移酶METTL1)確實是由RICK沉淀。典型的RBPs則存在于兩種方法的樣品中。
結果表明,RICK可分離出oligo(dT)捕獲研究中未鑒定的許多蛋白質(zhì),這些差異既不是由細胞培養(yǎng)、蛋白質(zhì)組學程序的變化引起的,也不是由EU標記引入的。
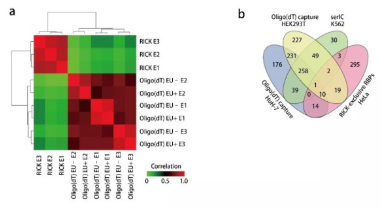
圖? 3
4)??RICK分離的蛋白質(zhì)的功能分析
對295個RICK獨特的RBPs進行GO分析,發(fā)現(xiàn)與有絲分裂有關的生物過程的富集(圖4a)。KEGG通路分析顯示前十個最顯著富集途徑中還包括“細胞周期”(圖4b)。

圖? 4
5)??RICK鑒定優(yōu)先結合非polyA RNAs的蛋白
首先研究者在標準RICK方法中添加了另一個步驟,即連續(xù)三輪與oligo(dT)包被的磁珠孵育以有效去除polyA RNAs,并通過RT-PCR得到驗證。LC-MS/MS鑒定出914種高置信度蛋白質(zhì),稱為’polyA耗盡的RICK蛋白質(zhì)’。其中,576種與標準RICK程序的720種高置信度蛋白質(zhì)重疊(圖5a)。進一步分析顯示295種RICK獨特RBPs中的204種(69.2%)與914種polyA耗盡的RICK蛋白質(zhì)重疊。另一方面,在不與RICK獨特的RBPs重疊的710種polyA耗盡的RICK蛋白中,563種存在于oligo(dT)捕獲數(shù)據(jù)集中(圖5b),說明這些蛋白具有結合polyA和非polyA RNA的混合傾向。western blotting證實polyA RNA去除之后RICK鑒定的RBPs(包括CDK1和METTL1)仍然富集。表明RICK可用于鑒定優(yōu)先結合非polyA RNAs的蛋白質(zhì)。
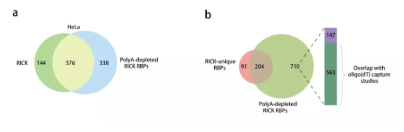
圖? 5
6)??與METTL1和CDK1相互作用的RNAs
研究者選擇了兩個RICK獨特的RBPs(METTL1和CDK1),使用PAR-CLIP測序研究與它們相互作用的RNAs。分析顯示METTL1與捕獲的RNAs廣泛結合,并且與tRNAs顯著結合(31.8%)。同時還存在對多種可能的非polyA RNAs的結合,例如內(nèi)含子RNAs(5.9%)和從基因間區(qū)域轉(zhuǎn)錄的RNAs(未注釋為lincRNAs;1.7%)(圖6)。另外,37.8%的序列被映射到mRNAs,但這些映射序列可能對應于未成熟轉(zhuǎn)錄本、circRNAs或ppRNAs。

圖? 6
接下來,通過RIP-qPCR發(fā)現(xiàn)METTL1與非polyA RNA VTRNA1-3相互作用。使用隨機六聚體引物可富集7種mRNAs,而使用oligo(dT)引物僅可相對擴增其中3種,支持了METTL1結合含有mRNA序列的非polyA RNAs(圖7d)。此外,分析METTL1的結合基序發(fā)現(xiàn)‘CUCUUCG’是2次重復實驗中最富集的結合基序(圖7e)。
對CDK1 PAR-CLIP測序數(shù)據(jù)進行分析得到相似結果。因此,METTL1和CDK1廣泛地與非polyA RNAs結合,暗示這兩種蛋白質(zhì)的RNA相關功能被忽視。
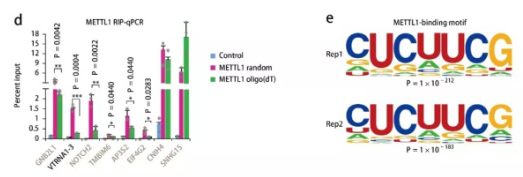
圖? 7
7)??RICK用于捕獲新生RNA相互作用組
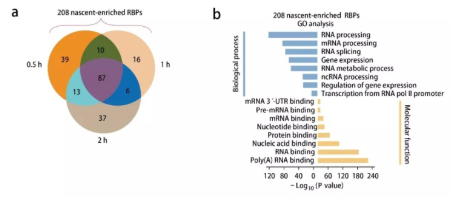
對HeLa細胞進行短時間EU標記(0.5、1和2 h),以研究RICK是否可以豐富捕獲與新生RNAs相互作用的蛋白質(zhì)。發(fā)現(xiàn)與16h RICK相比,由于摻入的EU較少,捕獲的RNAs和蛋白質(zhì)減少。短標記RICK中對應于5’UTR、CDS和3’UTR序列的百分比減少,表明成熟的mRNAs較少。
鑒定了208個不同的高置信度蛋白質(zhì)(0.5、1和2 h RICK分別鑒定149、119和143個高置信度蛋白質(zhì)),稱為新生富集的RBPs(圖8a)。208個新生富集的RBPs的GO和KEGG通路分析顯示了轉(zhuǎn)錄和RNA代謝過程的富集(圖8b)。進一步研究顯示,這208個新生富集的RBPs中只有43個未被oligo(dT)捕獲。這些發(fā)現(xiàn)表明新生的RNA轉(zhuǎn)錄
和局部代謝產(chǎn)物之間的聯(lián)系可調(diào)節(jié)表觀基因組和潛在的表觀轉(zhuǎn)錄組。
圖? 8
8) RICK捕獲mESCs的總RNA相互作用組
接下來,研究者旨在證明RICK可以應用于其他細胞類型。證實使用鏈霉親和素結合的辣根過氧化物酶在mESCs中可高效地摻入EU。鑒定出518個高置信度和304個低置信度蛋白質(zhì),其中358個專屬于RICK,稱為RICK獨有的mESC RBPs。358個RICK獨有的mESC RBPs中有95個蛋白在mESCs中的表達水平更高,指出其在mESC自我更新或多能性中的潛在作用。因此,RICK可以應用于除HeLa以外的不同細胞類型;需要進一步的研究來確定新鑒定的候選RBPs在mESCs自我更新/多能性中的作用(圖9)。

圖? 9


